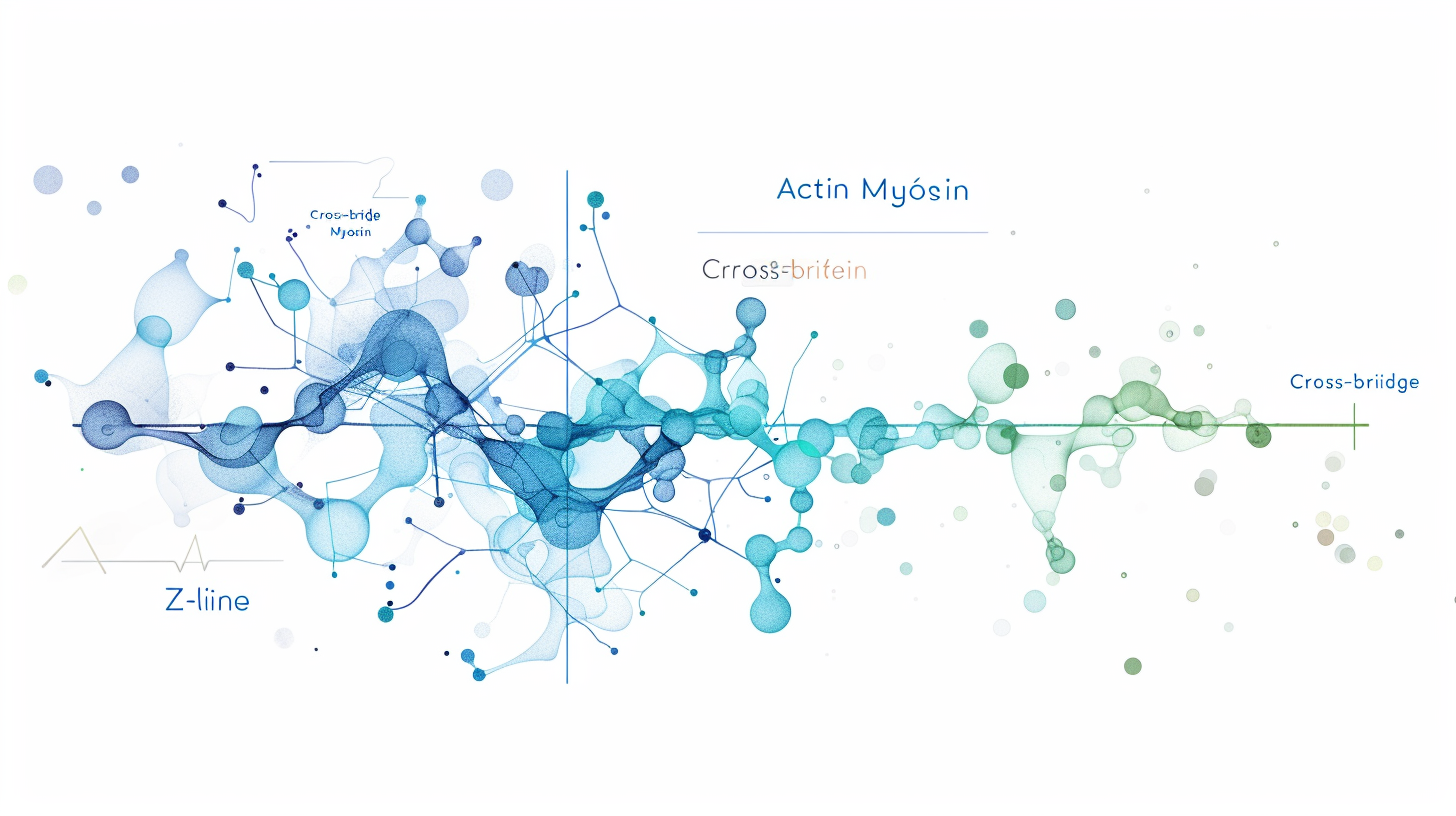
筋肉が縮む様子を、両手で綱を引っ張る動作に例えてみましょう。
- 太い綱(ミオシンフィラメント)が真ん中に固定されている
- 細い綱(アクチンフィラメント)が両側から伸びている
- ミオシンの「手」(クロスブリッジ)がアクチンをつかんで引き寄せる
- アクチンがミオシンの上を「滑るように」内側に引き込まれる
- 結果として、サルコメア全体が短くなる=筋肉が収縮する
重要なのは、フィラメント自体の長さは変わらないということです。アクチンがミオシンの上を滑ることで、全体が短くなります。これが「滑走説」の核心です。
語源
| 単語 | 語源 | 意味 |
|---|---|---|
| Sliding | 古英語 slīdan(滑る) | 滑走する |
| Filament | ラテン語 filamentum(糸) | 細い糸状の構造 |
| Theory | ギリシャ語 theoria(観察・考察) | 理論・学説 |
| Myosin | ギリシャ語 mys(筋肉) | ミオシン(太いフィラメント) |
| Actin | ギリシャ語 aktis(光線・放射) | アクチン(細いフィラメント) |
| Sarcomere | ギリシャ語 sarx(肉)+ meros(部分) | サルコメア(筋節) |
滑走説(Sliding Filament Theory)は1954年にHuxley & HansonおよびHuxley & Niedergerkeによって独立して提唱された、筋収縮のメカニズムを説明する最も基本的な理論です。
解説
筋肉の階層構造
滑走説を理解するには、まず筋肉の構造を把握する必要があります。
筋肉(Muscle)
└── 筋束(Fascicle)
└── 筋線維(Muscle Fiber)=筋細胞
└── 筋原線維(Myofibril)
└── サルコメア(Sarcomere)← 収縮の最小単位
├── アクチンフィラメント(細いフィラメント)
└── ミオシンフィラメント(太いフィラメント)サルコメアの構造
サルコメアはZ線からZ線までの1単位で、筋収縮の最小機能単位です。
| 構造 | 構成成分 | 役割 |
|---|---|---|
| Z線(Z-disc) | α-アクチニン | サルコメアの境界。アクチンが付着する。 |
| アクチンフィラメント | アクチン・トロポミオシン・トロポニン | 細いフィラメント。ミオシンが結合する部位を持つ。 |
| ミオシンフィラメント | ミオシン重鎖・軽鎖 | 太いフィラメント。クロスブリッジを形成する。 |
| A帯(A-band) | ミオシン全体(アクチン重複部含む) | 暗帯。ミオシンが存在する領域。収縮時も長さ不変。 |
| I帯(I-band) | アクチンのみの領域 | 明帯。収縮時に短くなる。 |
| H帯(H-zone) | ミオシンのみの領域 | 収縮時に短くなる・消失する。 |
| M線(M-line) | ミオメシン | ミオシンフィラメントを中央で固定する。 |
滑走説の7ステップ(クロスブリッジサイクル)
① 活動電位の発生
↓
② カルシウムイオン(Ca²⁺)の放出
↓
③ トロポニン・トロポミオシンの移動(結合部位の露出)
↓
④ クロスブリッジの形成(ミオシン頭部がアクチンに結合)
↓
⑤ パワーストローク(ミオシン頭部が傾き、アクチンを引き寄せる)
↓
⑥ ATPによるクロスブリッジの解離
↓
⑦ ミオシン頭部の再チャージ(ATPの加水分解)
↓
Ca²⁺が存在する限り①〜⑦を繰り返す各ステップの詳細
① 活動電位の発生 運動神経から筋線維に活動電位が伝わり、筋線維の細胞膜(筋鞘)を伝導します。T管(横行小管)を通じて筋線維内部まで届きます。
② カルシウムイオン(Ca²⁺)の放出 活動電位がT管を通じて筋小胞体(SR)を刺激し、Ca²⁺が筋形質(細胞質)に放出されます。安静時、Ca²⁺は筋小胞体に貯蔵されています。
③ トロポニン・トロポミオシンの移動 安静時、アクチンの結合部位はトロポミオシンによって覆われています(ブロック状態)。Ca²⁺がトロポニンに結合すると、トロポミオシンが横にずれ、アクチンの結合部位が露出します。
④ クロスブリッジの形成 露出したアクチンの結合部位にミオシン頭部が結合し、クロスブリッジが形成されます。この時点でミオシン頭部はADP+Piを持った「コックされた」状態です。
⑤ パワーストローク ADP+Piが放出される際にミオシン頭部が約45°傾き、アクチンフィラメントをZ線方向(内側)に約10nm引き寄せます。これが収縮力の発生源です。
⑥ クロスブリッジの解離 新しいATPがミオシン頭部に結合すると、ミオシンとアクチンの結合が解離します。ATPがなければ(死後硬直)この段階が起こらず、ミオシンとアクチンが結合したままになります。
⑦ ミオシン頭部の再チャージ ATPがATPaseにより加水分解されADP+Piになり、ミオシン頭部が元の「コックされた」高エネルギー状態に戻ります。Ca²⁺が存在する限りサイクルが繰り返されます。
収縮時の各帯の変化
| 部位 | 安静時 | 収縮時 |
|---|---|---|
| サルコメア全長 | 約2.0〜2.2μm(最適長さ) | 短くなる |
| A帯 | 一定 | 変化なし(ミオシン長さ不変) |
| I帯 | あり | 短くなる |
| H帯 | あり | 短くなる・消失 |
| フィラメント自体の長さ | 一定 | 変化なし |
重要:フィラメント自体は縮まない。アクチンがミオシンの上を滑ることでサルコメアが短くなる。
弛緩のメカニズム
神経刺激が止まると以下が起こります。
- 筋小胞体がCa²⁺を能動輸送で回収する(ATPを消費)
- 細胞質のCa²⁺濃度が低下する
- トロポミオシンがアクチンの結合部位を再び覆う
- クロスブリッジが形成されなくなる
- 筋肉が弛緩する
弛緩もATPを消費する能動的なプロセスです。
豆知識
死後硬直(Rigor Mortis)と滑走説
死後硬直は滑走説で完璧に説明できます。
- 死後、ATPの産生が停止する
- Ca²⁺が筋小胞体から漏れ出し、筋形質に拡散する
- クロスブリッジが形成される
- しかしATPがないためミオシンがアクチンから解離できない
- 結果として筋肉が硬直した状態で固まる
これが「ATPは収縮だけでなく弛緩にも必要」という証拠です。
サルコメアの最適長さとトレーニング
サルコメアが最大の力を発揮できる長さは約2.0〜2.2μmです。これは「長さ-張力関係(Length-Tension Relationship)」と呼ばれる概念です。
- 短すぎる(過収縮):アクチンフィラメントが重なり合い、クロスブリッジが形成できない
- 最適長さ:クロスブリッジが最大数形成できる
- 長すぎる(過伸展):アクチンとミオシンの重複が少なくなり、クロスブリッジ数が減る
これがストレッチポジションでの筋肥大(エキセントリックトレーニング)の理論的根拠の一つです。
「速筋と遅筋でクロスブリッジサイクルが違う」
ミオシン重鎖(MHC)にはアイソフォームがあり、サイクルの速度が異なります。
| 筋線維タイプ | ミオシンATPase活性 | クロスブリッジサイクル速度 |
|---|---|---|
| タイプI(遅筋) | 低い | 遅い(持久力向き) |
| タイプIIa | 中程度 | 中程度 |
| タイプIIx(速筋) | 高い | 速い(瞬発力向き) |
関連論文
| 著者・年 | 内容 | 主な結論 |
|---|---|---|
| Huxley & Hanson (1954) | 滑走説の原著論文(Nature誌) | 電子顕微鏡観察により、筋収縮時にA帯長は変化せずI帯・H帯が短縮することを発見。滑走説を提唱。 |
| Huxley & Niedergerke (1954) | 滑走説の同時独立提唱(Nature誌) | 干渉顕微鏡を使用した観察により同様の結論を独立して発表。 |
| Gordon et al. (1966) | 長さ-張力関係の実証 | サルコメア長と筋収縮力の関係を実験的に示し、最適長さ(2.0〜2.2μm)を特定。 |
| Schoenfeld (2010) | 筋肥大のメカニズムレビュー | 機械的張力・代謝ストレス・筋損傷が筋肥大の三大刺激。滑走説に基づくクロスブリッジ理論が基盤。 |
| McMahon et al. (2021) | ストレッチポジションでの筋肥大 | エキセントリック局面(ストレッチポジション)でのトレーニングが筋肥大を促進する可能性を示唆。 |
よくある質問
- Q滑走説でフィラメント自体は縮みますか?
- A
縮みません。アクチンフィラメントもミオシンフィラメントも、収縮中に長さは変化しません。アクチンがミオシンの上を滑り込むことでサルコメア全体が短くなります。これが「滑走」説の核心です。
- QATPは筋収縮にどう使われますか?
- A
ATPは3つの場面で使われます。①クロスブリッジの解離(ミオシンとアクチンを引き離す)、②ミオシン頭部の再チャージ(ADP+Piへの加水分解)、③弛緩時のCa²⁺の筋小胞体への回収です。ATPがなければ収縮も弛緩もできません。
- Qカルシウムイオン(Ca²⁺)はなぜ重要ですか?
- A
安静時はトロポミオシンがアクチンの結合部位をブロックしています。Ca²⁺がトロポニンに結合することでトロポミオシンが移動し、ブロックが解除されてクロスブリッジが形成できるようになります。Ca²⁺は筋収縮の「スイッチ」です。
- Q筋肉痛と滑走説の関係は?
- A
筋肉痛(DOMS)は主にエキセントリック収縮(アクチンとミオシンが引き離される方向の力)による筋線維・結合組織の微細損傷が原因とされています。過度のクロスブリッジへの負荷がサルコメアレベルでの損傷を引き起こすと考えられています。
- Qトロポニンとトロポミオシンの違いは何ですか?
- A
トロポミオシンはアクチンフィラメントに沿って走る細長いタンパク質で、安静時にアクチンの結合部位を物理的にブロックします。トロポニンはトロポミオシンに結合するタンパク質複合体で、Ca²⁺を受け取るセンサーです。Ca²⁺を受け取ったトロポニンがトロポミオシンを動かします。
- Q最大筋力を発揮するにはサルコメアの長さが関係しますか?
- A
はい。サルコメアが最適長さ(2.0〜2.2μm)のとき、アクチンとミオシンの重複が最大となりクロスブリッジが最も多く形成されるため、最大の力を発揮できます。筋肉が過度に短縮または伸張している状態では力が低下します。
- Q滑走説は今も正しいのですか?
- A
基本的な枠組みは1954年の提唱から現在まで支持され続けています。ただし分子レベルの詳細(ミオシン頭部の動きの精密な機構など)は現代の技術で更新・精緻化されています。「ワーキングストロークモデル」など発展的理論も提唱されていますが、滑走説の根幹は変わっていません。
理解度チェック
問題1:滑走説において、筋収縮時に長さが変化しないものはどれか?
① サルコメア全体
② I帯
③ H帯
④ アクチン・ミオシンフィラメント自体
正解:④ 解説:滑走説の最重要ポイントです。フィラメント自体は縮まず、アクチンがミオシンの上を滑ることでサルコメアが短くなります。A帯の長さも変化しません。
問題2:安静時にアクチンの結合部位をブロックしているタンパク質はどれか?
① トロポニン
② トロポミオシン
③ アクチニン
④ チチン
正解:② 解説:安静時はトロポミオシンがアクチンの結合部位を覆っています。Ca²⁺がトロポニンに結合することでトロポミオシンが移動し、結合部位が露出します。
問題3:クロスブリッジサイクルでATPが直接関与するステップはどれか?
① パワーストロークの発生
② Ca²⁺の筋小胞体への回収
③ クロスブリッジの形成
④ トロポニンへのCa²⁺結合
正解:② 解説:ATPは①クロスブリッジの解離、②ミオシン頭部の再チャージ、③Ca²⁺の筋小胞体への能動輸送(弛緩)に使われます。パワーストロークはADP+Piの放出時に発生します。
問題4:死後硬直(Rigor Mortis)が起こる直接の原因はどれか?
① Ca²⁺が筋小胞体に回収されすぎるから
② ATPがなくなりクロスブリッジが解離できないから
③ ミオシンフィラメントが変性するから
④ トロポニンがCa²⁺を放出できなくなるから
正解:② 解説:死後ATPが枯渇するとミオシンとアクチンの結合を解離できなくなり、クロスブリッジが固定されたまま硬直します。これは「弛緩にもATPが必要」という証拠です。
問題5:サルコメアの最適長さとして正しいものはどれか?
① 約0.5〜1.0μm
② 約2.0〜2.2μm
③ 約4.0〜5.0μm
④ 約10μm以上
正解:② 解説:サルコメアの最適長さは約2.0〜2.2μmです。この長さでアクチンとミオシンの重複が最大となり、最も多くのクロスブリッジが形成されて最大の力が発揮されます。
問題6:速筋線維(タイプIIx)と遅筋線維(タイプI)のクロスブリッジサイクルの違いとして正しいものはどれか?
① タイプIのほうがクロスブリッジサイクルが速い
② タイプIIxのほうがミオシンATPase活性が高くサイクルが速い
③ 両者のサイクル速度に差はない
④ タイプIIxはクロスブリッジを形成しない
正解:② 解説:タイプIIxはミオシン重鎖(MHC)のアイソフォームが異なり、ATPase活性が高いためクロスブリッジサイクルが速く、瞬発的な力発揮が得意です。タイプIは逆に遅く、持久力向きです。
問題7:筋収縮時にI帯とH帯はどうなるか?
① I帯が長くなり、H帯も長くなる
② I帯が短くなり、H帯も短くなる(消失することもある)
③ I帯は変化せず、H帯のみ短くなる
④ I帯・H帯ともに変化しない
正解:② 解説:収縮時にアクチンがミオシン上を滑り込むことで、アクチンのみの領域(I帯)とミオシンのみの領域(H帯)が短くなります。A帯(ミオシン全体の領域)のみ変化しません。
覚え方
【語呂合わせ:クロスブリッジサイクルの順番】
「活動・カル・トロ・クロ・パワー・解離・再チャージ」
活動(活動電位)→ カル(Ca²⁺放出)→ トロ(トロポミオシン移動)→ クロ(クロスブリッジ形成)→ パワー(パワーストローク)→ 解離(ATPでミオシン解離)→ 再チャージ(ミオシン頭部リセット)
【収縮時の帯の変化を覚える】
「A帯は変わらない。I帯とH帯は短くなる。フィラメントは縮まない。」
A → Always same(常に同じ) I → Inevitably shorter(必ず短くなる) H → Has to shrink(縮まなければならない)
【Ca²⁺の役割をひとことで】
「Ca²⁺=筋収縮のスイッチ。Ca²⁺が来たらON、去ったらOFF」
トロポニン(センサー)がCa²⁺を感知 → トロポミオシン(蓋)が開く → アクチン(結合部位)が露出 → ミオシン(手)がつかむ
まとめ
- 滑走説の核心:筋肉が縮む時、アクチンとミオシンフィラメント自体の長さは変わらない。
- アクチンがミオシンの上を滑ることでサルコメアが短くなる。
- Ca²⁺がスイッチ:安静時はトロポミオシンがアクチンの結合部位をブロック。Ca²⁺がトロポニンに結合することで初めてクロスブリッジが形成できる。
- ATPは収縮にも弛緩にも必要:クロスブリッジの解離・ミオシン頭部の再チャージ・Ca²⁺の回収すべてにATPが必要。死後硬直はATP枯渇の証明。
- A帯は変化しない:収縮時にI帯・H帯は短くなるが、A帯(ミオシンの存在領域)の長さは変化しない。これが滑走説の物理的証拠。
- 最適長さ2.0〜2.2μm:サルコメアがこの長さのとき最大の力を発揮。ストレッチポジションでのトレーニングの理論的根拠のひとつ。
必須用語リスト
| 用語 | 意味 |
|---|---|
| 滑走説(Sliding Filament Theory) | 筋収縮時にアクチンがミオシン上を滑ることでサルコメアが短くなるという理論。1954年提唱。 |
| サルコメア | 筋収縮の最小機能単位。Z線からZ線まで。安静時の最適長さは2.0〜2.2μm。 |
| アクチンフィラメント | 細いフィラメント。トロポニン・トロポミオシンと複合体を形成する。 |
| ミオシンフィラメント | 太いフィラメント。クロスブリッジ(頭部)を持ち、アクチンに結合して力を発生させる。 |
| クロスブリッジ | ミオシン頭部がアクチンに結合した状態。力発生の最小単位。 |
| パワーストローク | ミオシン頭部が傾いてアクチンを引き寄せる動作。約10nmの移動。ADP+Pi放出時に発生。 |
| トロポニン | アクチンフィラメント上のCa²⁺センサータンパク質。Ca²⁺結合でトロポミオシンを移動させる。 |
| トロポミオシン | 安静時にアクチンの結合部位をブロックするタンパク質。Ca²⁺シグナルで移動し結合部位を露出する。 |
| 筋小胞体(SR) | Ca²⁺を貯蔵・放出する筋線維内の膜構造。活動電位に応答してCa²⁺を放出する。 |
| T管(横行小管) | 活動電位を筋線維内部に伝導する管状構造。筋小胞体と連絡する。 |
| A帯 | ミオシンフィラメントが存在する領域(暗帯)。収縮時も長さは変化しない。 |
| I帯 | アクチンフィラメントのみの領域(明帯)。収縮時に短くなる。 |
| H帯 | ミオシンフィラメントのみの領域。収縮時に短くなり、完全収縮時に消失する。 |
| 長さ-張力関係 | サルコメアの長さと発揮できる筋力の関係。最適長さ(2.0〜2.2μm)で最大力を発揮。 |
| 死後硬直(Rigor Mortis) | 死後ATPが枯渇しクロスブリッジが解離できなくなった状態。弛緩にもATPが必要な証拠。 |
コメント